




Request a free consultation with one of our experts and a member of our team will contact you within 24 hours.
Complete Request FormOur Student Success Team is available every day to answer all your questions.
Chat Now or call 888-427-7737With a highly customizable study planner tool, Live Review Sessions 6 days/week at no extra charge, plus practice tests, hundreds of on-demand video explanations, you can truly tailor your MCAT preparation.
Blueprint MCAT teaches you how the MCAT thinks. Our exams & interface are the most representative, after the AAMC, so you will be extra prepared and confident on test day.
Our Self-Paced Course was created by 524+ scoring instructors & more than 50 of our tutors have scored 520+ on the MCAT. Meet our MCAT instructors and tutors.
Because we’re so confident in our methodology, we guarantee that your MCAT exam score will increase or your money back. Recent results showed future medical students increased their MCAT score by an average of 15 points.
Check out how Blueprint courses
helped future medical students like you achieve their MCAT score goals
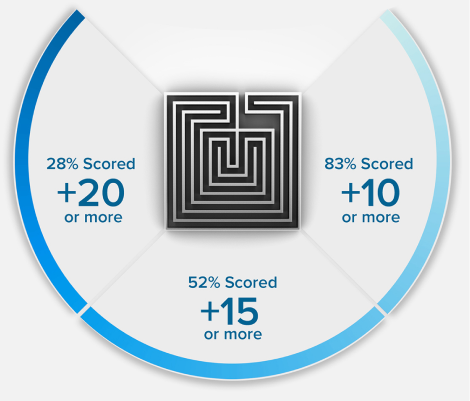
15 POINTS? WE DON'T STOP THERE.
+15
POINTS ON AVERAGE*
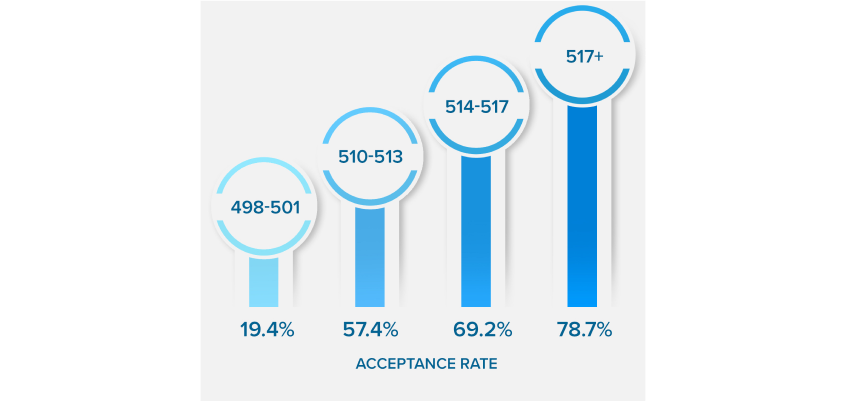
NEARLY QUADRUPLE
YOUR ACCEPTANCE RATE
*Results based on our Live Online MCAT Course Cohort. See more details on score increases.
Introducing Blue
Get the “why” behind right and wrong answers and transform your approach with a 24/7 tutor trained by MCAT experts and totally personalized to you.
“Even if I didn’t fully know how to phrase my question, it seemed to understand what I was asking and it gave a great answer!”
- Thomas G., Real AI Tutor User

Blueprint instructors aren't just brilliant MCAT experts and content masters, they're also the best teachers around with years of experience under their belt. Teaching is a skill, and we only hire instructors that have shown the ability to take the most advanced science concepts and explain them in easy to understand terms — and make it as fun as possible!
You will be taken to another Blueprint Prep company site or one of our partners. Do you want to continue?
Request a consultation with an MCAT Advisor, or call 888-530-6398 for immediate assistance.